
1. Predator-prey dynamics in biological control
Biological control of pests using natural enemies is becoming increasingly used in pest control. In addition to the applied component, predator-prey dynamics in biological control can contribute to theory on the dynamics of coupled predator prey systems. Applied contributions to biological control are often focused on the national level, however benefits of biological control at the national level can potentially be beneficial internationally, as many pest species are invasive and occur worldwide (e.g. Western flower thrips (Frankliniella occidentalis)). To this end identifying suitable natural enemies to control pest species not only provides insight to test and extend current theory on predator-prey systems but opens a much-needed applied component which can be drafted into policy. Currently I, in collaboration with Dr. Arne Janssen and Dr. Paul van Rijn, am investigating the role of alternative food in biological control of dry bulb mites and Western flower thrips. Alternative food in a predator-prey system ensures populations of predators are maintained in the absence of prey. A key aspect of the role of alternative food in biological control that is currently lacking is the quality and frequency of application. Using data from several laboratory and greenhouse experiments we construct detailed population-dynamical models which incorporate an alternative food supply. Outcomes can be directly applied to biological control methods but can also contribute theoretically to coupled predator-prey systems.

Figure shows the impact of the addition of alternative food to a system. (A) SImulations of the change in pest density over time; (B) changes in predator density; (C) change in predator satiation (effectively how full the predator is). Black line indicates no alternative food added, purple line indicates alternative food of low quality, yellow line alternative food of high quality and red line when predators are absent from the system. Dashed line in (C) indicates maintenance level threshold.
2. Changes in population dynamics through dispersal and variable environments
Individual and population level effects of dispersal
One of my model systems is the bulb mite ( Rhizoglyphus robini) and the theoretical framework that I use is a form of discrete structured-time modelling (Integral Projection Models, deterministic and stochastic). In population models, dispersal is often considered as a population-level process, however the focus has shifted from the population level to the level of individual organisms. As its the behaviour and demographic changes of individuals that shape the dynamics of populations, to fully understand the effects of dispersal on natural systems it is necessary to incorporate individual level differences in dispersal behaviour in population models. In collaboration with Dr. Isabel Smallegange, the aim is to determine the influence of dispersal propensity on population size and structure, life histories and how these change with stochastic environments. I am also collaborating with Dr. Gregory Roth to develop a model that incorporates the costs of dispersal life-histories, emigration effects and immigration effects on natal populations.
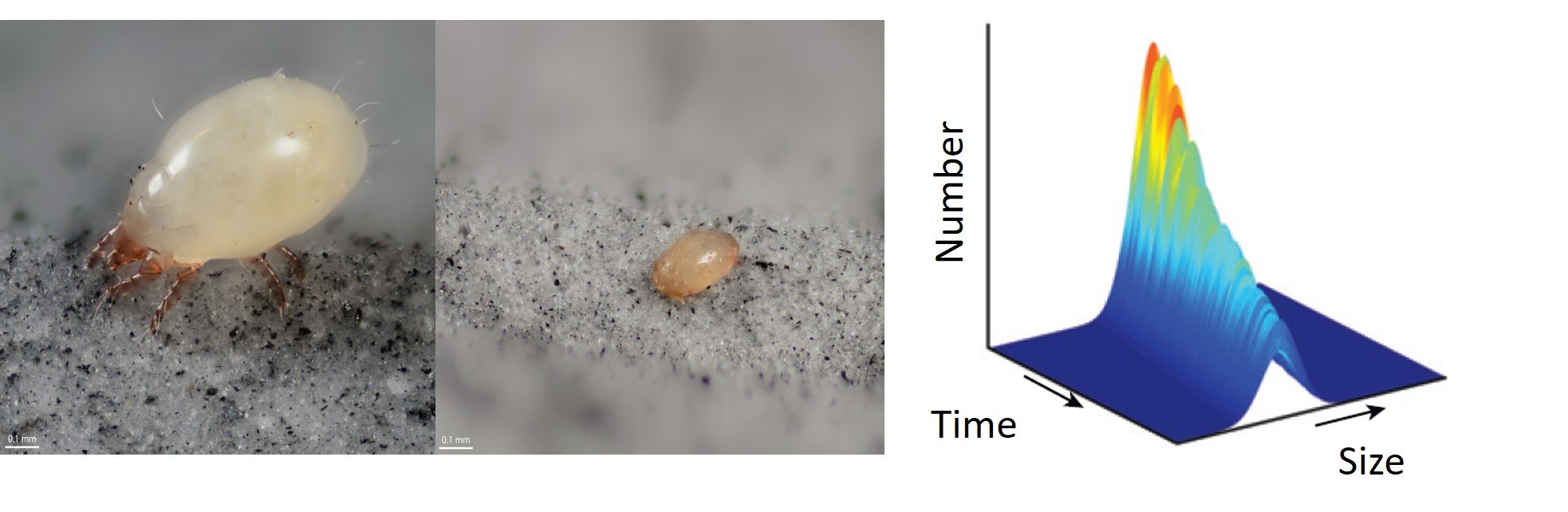
Left panel: Adult female bulb mite (left), juvenile dispersal stage (deutonymph) of the bulb mite (right). Right panel: graph showing an example of how a size distribution can change over time due to changes in births and deaths of individuals of different sizes.
Mechanisms of eco-evolutionary dynamics
Given that the feedbacks between ecology and contemporary evolution are now common place, there is a need to understand what the influence of these feedbacks are at various levels of biological understanding, as well as how the feedbacks are influenced by changing environments. I focus on the individual and population level in an attempt to understand how various life-history traits may be affected and how they in turn may affect eco-evolutionary feedback - this requires a mechanistic understanding. To do so I, in collaboration with Dr. Isabel Smallegange, investigate how individuals with different life-histories influence population dynamics and structure and how these population changes in turn affect selection pressures, and ultimately trait change, within the population.

Figure shows how a change in the environment (here harvesting of the two different male morphs - fighter male (FM) or scrambler male (SM)) impacts: A) probability an individual will develop into a fighter; B) probability an individual will develop into a fighter male given the size of the final juvenile development stage (Quiescent tritonymph); C) adult and quiescent tritonymph body size of females and the two male morphs.
Impact of calorie restriction (CR) on longevity and other life-history traits
CR theory predicts prolonged life expectancy due to restricted food intake. However, there is some controversy over the benefits of CR. Furthermore, interaction effects are not well established, with most CR research largely neglecting interaction effects. In collaboration with Dr. Roberto Salguero-Gómez, I investigate, experimentally, the impacts of several interacting factors (food quality and timing) in addition to CR on longevity in long-lived planarian species, Schmidtea mediterranea and Dugesia tahitiensis. From the data generated by the experiments we aim to construct population models in an effort to understand the impacts of CR and interaction effects on the population dynamics of the two species
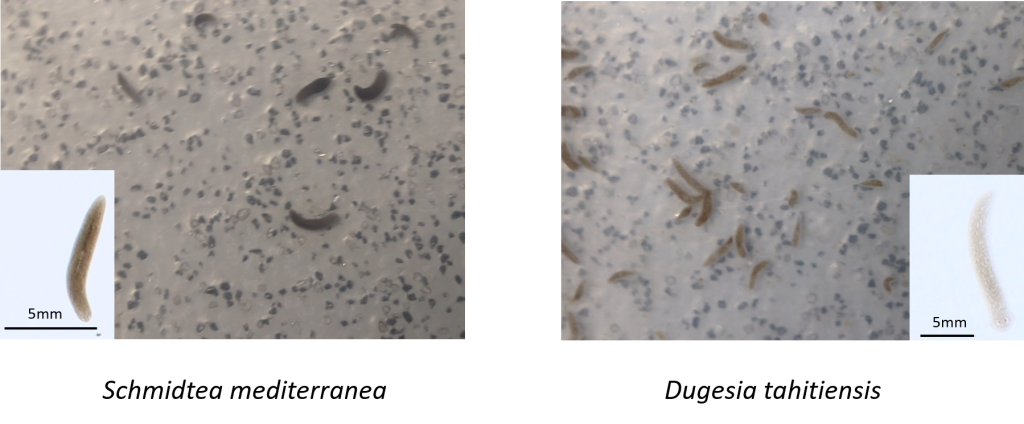
The two planarian species used in our research, note the differences in size. Schmidtea mediterranea (left) and Dugesia tahitiensis (right).